LE
PIANTE, FLUSSO E FUNZIONI DALLE RADICI ALLE FOGLIE – da enciclopedia Treccani
Acqua
nelle piante
Nelle piante non c'è nessuna struttura che spinge l'acqua
fino alle foglie dei rami più alti e sono ancora le caratteristiche delle
molecole di acqua che permettono il trasporto delle sostanze dalle foglie alle
radici e dalle radici alle foglie.
Nelle piante superiori, si sono sviluppati particolari
tessuti di conduzione, ovvero delle strutture
attraverso le quali l'acqua può raggiungere le foglie più alte e distribuire a
tutte le parti della pianta le sostanze organiche sintetizzate nelle foglie.
Anche nelle piante non vascolari il trasporto avviene nei due sensi, ma i
tessuti di conduzione sono meno differenziati.
Negli alberi ad alto fusto, come ad esempio le esemplari
sequoie, l'acqua deve percorrere distanze che possono raggiungere anche diverse
decine di metri. I meccanismi che contribuiscono alla risalita dell'acqua sono
diversi, e ancora una volta ricollegabili alle sue caratteristiche
microscopiche.
L'acqua del terreno, infatti, passa per osmosi all'interno
dei peli radicali attraverso la membrana costituita dalle cellule epidermiche.
I peli radicali, quindi, assumono dal terreno soluzioni
di ioni inorganici e di piccole molecole di sali minerali, permettendo
l'instaurarsi di un flusso mantenuto dalle differenze di concentrazione tra
l'esterno (soluzioni poco concentrate nel terreno) e l'interno della pianta
(soluzioni molto concentrate nelle cellule). Si genera così quella che viene definita 'pressione radicale' che può essere
sufficiente a spingere l'acqua fino ad altezze modeste, ma che non è certo in
grado di farle raggiungere le sommità di grossi alberi o dei lunghi steli delle
piante rampicanti. La pressione richiesta in questi casi è molto ingente e di gran lunga più elevata di quella sviluppata per osmosi,
né il fenomeno può essere spiegato con l'aspirazione: anche il vuoto assoluto
all'interno dell'albero non potrebbe sollevare una colonna d'acqua a più di
Dati i minuscoli diametri dei vasi xilematici in cui
scorrono acqua e sali minerali, le forze di capillarità possono svolgere un
ruolo importante e, affiancandosi alla pressione radicale, contribuire a
potenziare l'ascesa della linfa. Le caratteristiche polari delle molecole di
cellulosa, costituente principale delle pareti cellulari delle piante, rendono
questa sostanza altamente idrofilica
e in grado, quindi, di instaurare forze di adesione particolarmente intense con
le molecole d'acqua.
Ma anche prendendo in considerazione la capillarità
non si riesce a dar conto della risalita delle molecole d'acqua fino a quote
considerevoli.
C'e bisogno di un ulteriore forza trainante che faccia
arrivare la linfa fino alla sommità della chioma. Così le piante sfruttano
attivamente la perdita d'acqua che ha luogo a livello delle foglie, traendo
vantaggio dalla tendenza alla disidratazione che rappresenterebbe un potenziale
problema. Proprio dalla superficie delle foglie esposte all'aria
l'acqua evapora continuamente attraverso particolari aperture: gli stomi.
Questo fenomeno, chiamato traspirazione viene regolato
dalla maggiore o minore apertura degli stomi stessi: se sono aperti l'acqua
lascia la foglia in forma di vapore, se sono chiusi, l'acqua rimane trattenuta
al suo interno e si evitano perdite eccessive di liquido.
Il passaggio delle molecole dalla foglia all'atmosfera nel
corso della traspirazione crea, a causa del grandissimo grado di coesione
interno, una situazione di squilibro e di 'tensione'
che concretamente risulta in una forza diretta verso l'alto che attrae l'acqua
nella parte alta del sistema vascolare, vincendo la forza di gravità. Così il
flusso d'acqua, trascinato dalle molecole che stanno evaporando, scorre senza
frammentarsi dalla base dell'albero alla chioma.
L'acqua resiste a questa tensione come un elastico teso che
si deforma ma non si rompe a causa della fitta rete di
legami idrogeno che la mantiene coesa. Il mantenimento di una colonna continua
di liquido è fondamentale per la sopravvivenza della pianta: un'interruzione
dovuta, per esempio, alla presenza di bolle che ostruissero il passaggio,
provocherebbe la separazione in due spezzoni della colonna stessa con la parte
inferiore che non potrebbe più essere richiamata verso
Pressione radicale e traspirazione cooperano alla salita
dell'acqua fino alle foglie: l'una spinge dal basso, l'altra tira dall'alto.
Dalle radici che forniscono costantemente nuova soluzione salina fino ai
germogli apicali, la pianta è quindi percorsa da un flusso continuo di acqua
che, attraverso la traspirazione, si disperde nell'ambiente. Questo fenomeno ha
delle dimensioni difficilmente immaginabili: un modesto albero di betulla
traspira più di trecento litri d'acqua al giorno,
molta più di quella che evaporerebbe dal suolo nudo; così anche una semplice pianta
di grano, che pesa meno di mezzo chilo al momento del raccolto, ha assunto,
trasportato e liberato nell'atmosfera quasi due quintali di acqua durante la
sua vita.
Fotosintesi
e respirazione
Nel tessuto verde delle foglie avviene la fotosintesi che
comprende a) i complicati processi endoergonici
relativi all'attivazione e al trasferimento degli elettroni che dall'acqua
vanno a ridurre l'accettore NADP + mentre si libera ossigeno molecolare
nell'ambiente e b) le successive reazioni di organicazione
del carbonio in cui la CO2 viene ridotta a
carboidrato. I reagenti chimici sono dunque l'acqua che arriva alle foglie
dalle radici e l'anidride carbonica che dall'aria entra nelle foglie attraverso
gli stomi.
Nelle foglie e nei fusti, in parallelo allo xilema, si
trovano i tubi del floema, (comunemente detti vasi cribrosi) in cui avviene il
trasporto delle sostanze costruite con la fotosintesi fino alle varie parti
della pianta.
Mentre l'acqua sale nello xilema per ragioni fisiche, sono
i tubi del floema a mettere in comunicazione i luoghi di produzione e quelli di
consumo che utilizzano di solito la sorgente più vicina. La soluzione con i
prodotti della fotosintesi (linfa elaborata) deve essere distribuita in tutte
le direzioni, e per questo servono i meccanismi del trasporto attivo che
impiegano altra energia chimica.
Le sostanze vegetali, formate a partire
dagli zuccheri, sono alimento indispensabile per gli animali incapaci di
sfruttare altre forme di energia e di costruire da soli le sostanze necessarie
alla loro esistenza.
Quando gli animali si nutrono e respirano, all'interno
delle loro cellule (nei mitocondri), gli zuccheri (assunti con la nutrizione) e
l'ossigeno (assunto con la respirazione) vengono
reciprocamente trasformati. In presenza di ossigeno,
infatti, circa il 50% della materia organica prodotta con la fotosintesi viene
usata come combustibile per la respirazione cellulare e complessivamente
ossidata ad anidride carbonica, liberando circa 686 kcal per mole di glucosio,
con un rendimento di circa il 60%. Una parte dell'energia chimica rimasta nelle
molecole fotosintetizzate viene
trasferita ad altre molecole di ATP che si formano nel processo respiratorio,
circa 32 per ogni molecola di zucchero: l'ATP, trasportato nei luoghi
appropriati della cellula, servirà per il movimento, il calore e la vita
dell'organismo.
Al termine del processo respiratorio, l'accettore finale
degli elettroni e degli ioni idrogeno che componevano la molecola dello
zucchero è proprio l'ossigeno, fortemente elettronegativo,
che viene così ridotto ad acqua. Contemporaneamente, il carbonio si ossida a CO2,
e viene eliminato nell'ambiente con i meccanismi
respiratori.
Si ricostruiscono così, con una sorta di demolizione
ossidativa degli zuccheri che coinvolge processi di fosforilazione e di
produzione di ATP, le due molecole di acqua e di anidride carbonica utilizzate nella fotosintesi.
Mentre fotosintesi e respirazione sono due processi che dal
punto di vista della materia procedono in senso inverso e che - di conseguenza
- fanno parte di un ciclo che si avvia e si conclude
con acqua e anidride carbonica, considerazioni analoghe non si possono fare per
l'energia che non ha una 'storia' ciclica, ma viene degradata nei processi
vitali. L'energia solare, infatti, attraverso le sue successive trasformazioni
in altre forme di energia (chimica, meccanica, ecc.) ha reso possibile il
funzionamento dei viventi e infine, dissipandosi completamente in calore, si è
dispersa nell'Universo - cosa che sarebbe successa ugualmente
anche se i viventi non l'avessero per un momento bloccata e trasformata
per il loro mantenimento.
Tessuti Vegetali: i Tessuti Conduttori – da
giardinaggio.efiori.com
\"")
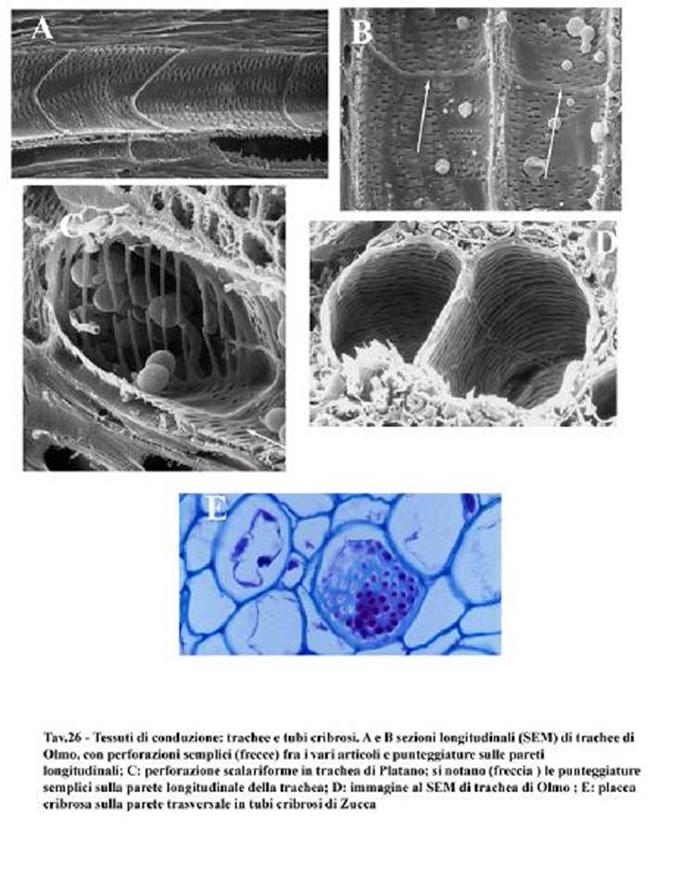
I tessuti
conduttori sono costituiti da lunghe file di cellule che si sovrappongono e, in
questa sovrapposizione dove l'una è a contatto con l'altra, una parte o si
perfora o viene modificata e va a formare una sorta di
tubo molto lungo. Sono i vasi,
quella struttura, cioè, preposta al collegamento
dell'apparato radicale con la parte aerea di una pianta.
Nelle piante di una certa età, anche se di
notevoli dimensioni e diametro del fusto, i vasi si trovano pochi millimetri
sotto la corteccia.
In questo poco spazio, dunque, avviene la quasi totalità del trasporto della linfa fra chioma e radici. Sono
tessuti complessi essendo costituiti da diversi tipi di cellule, raggruppati in
fasci insieme
alle fibre e
si dividono in vasi
xilematici o legnosi
che formano lo xilema
o legno e in vasi floematici o
cribrosi che
formano il floema
o libro.I
primi (vasi xilematici)
hanno il compito di trasportare la linfa
grezza (composta essenzialmente da acqua e sali
minerali) dalle radici alle foglie e sono vasi costituiti da
cellule morte (trachee
e tracheidi)
con protoplasma assente, a parete
|
|
ispessita e, in parte,
lignificata. Si possono quindi paragonare a dei tubicini come fossero tante
cannucce e, dato che in essi la linfa viene
"aspirata o succhiata" (che dir
si voglia) dall'alto tramite l'evaporazione che avviene nella parte aerea (il
meccanismo, in pratica, della cannuccia e della bibita) possiamo dire che
esiste una sorta di tensione
interna alla pianta perchè ciò avvenga
(incredibili piante! Non è cosa da poco vincere la forza di gravità eppure
loro...). Questa tensione interna potrebbe esporre i
vasi al collassamento ma proprio la lignificazione
di una parte della parete del vaso, impedisce che
ciò accada. |

{kind=link}
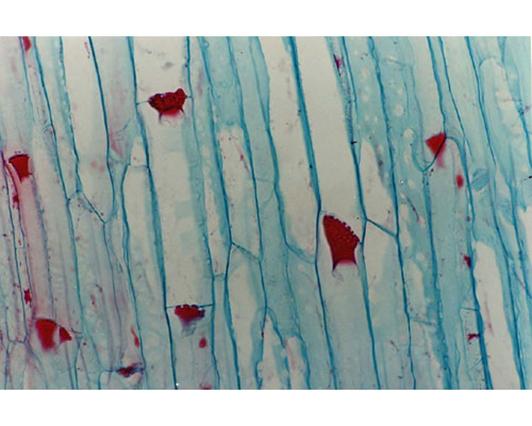
I vasi floematici, invece, trasportano la linfa elaborata (costituita da una soluzione acquosa in cui sono contenute sostanze organiche prodotte dalla fotosintesi -gli zuccheri- ormoni e aminoacidi), dalle foglie a tutto il resto della pianta e alle radici e sono costituiti da cellule vive comunque modificate. In questi vasi il trasporto della linfa avviene grazie ad una sorta di pressione che "spinge" verso il basso. Le pareti di questi vasi, sono ispessite ma mai lignificate.
{kind=link}
In aggiunta alle strutture preposte alla conduzione, nel legno e nel libro (xilema e floema) si trovano anche fibre e cellule parenchimatiche.
Dicevamo che trachee
e tracheidi formano lo xilema o legno,
disposto nella parte più centrale della pianta, mentre i vasi cribrosi
costituiscono il libro o floema che si trova
nella parte più interna della corteccia. Le trachee (vasi aperti), a
differenza delle tracheidi, sono maggiormente efficienti nel trasporto della
linfa grezza delle piante più evolute come ad esempio le latifoglie
mentre le tracheidi (detti vasi chiusi perchè la
linfa deve sempre attraversare la lamella mediana per poter passare) sono
presenti in piante più primitive come le conifere e, per la loro
conformazione più irregolare, per la sezione più stretta, presentano più
ostacoli al passaggio della linfa. 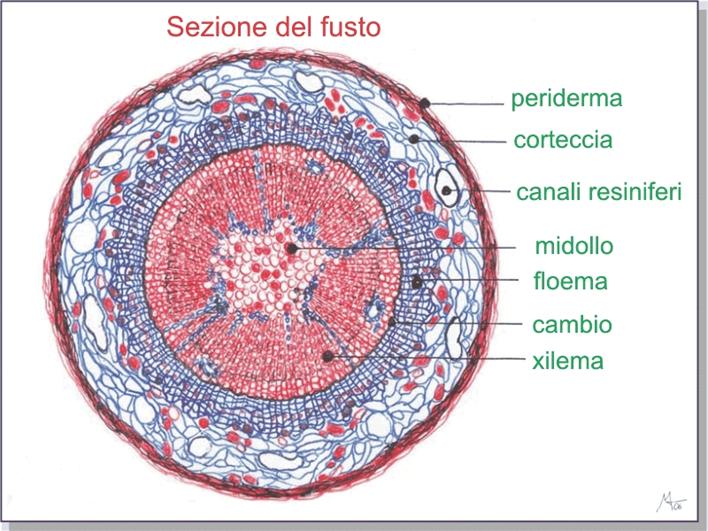
{kind=link}
Le fibre, invece, hanno la funzione di aumentare la resistenza dei fasci a fronte delle sollecitazioni di cui parlavamo prima mentre le cellule vive sono deputate soprattutto alla regolazione del flusso della linfa grezza all'interno di trachee e tracheidi. In autunno, nelle caducifolia, le cellule vive danno origine a delle estroflessioni somiglianti a piccolissime ernie del tessuto per fermare la circolazione della linfa, chiamate tille. Con la bella stagione, invece, quando sarà indispensabile far ripartire il flusso della linfa grezza, le tille verranno riassorbite.
{kind=link}
Le cellule vive hanno anche un'altra funzione nei mesi invernali, sui vasi cribosi (floematici) cioè quella di bloccarne l'attività "incrostando" le zone di perforazione tra cellula e cellula da cui percola la linfa elaborata, in maniera tale da impedirne il flusso. Anche queste "incrostazioni" verranno rimosse, poi, con l'approssimarsi della bella stagione.
Ma oltre al collegamento verticale all'interno della pianta, atto a garantire la distribuzione della linfa dalle radici alla chioma e viceversa, esiste anche il collegamento trasversale tra l'interno e l'esterno ed è garantito dai raggi midollari o parenchimatici, costituiti da una serie di cellule allungate e disposte in fila e che, oltre ad avere la funzione di conduzione delle sostanze nutritive hanno anche quella di costituire una riserva di amidi e sostanze minerali.